



Βίντεο Νο 379

![Η ΜΝΗΜΗ ΤΟΥ ΝΕΡΟΥ Νο3 (ΛΥΚ ΜΟΝΤΑΝΙΕ)| ΝΤΟΚΙΜΑΝΤΕΡ ΣΤΟ ΓΑΛΛΙΚΟ ΚΑΝΑΛΙ FRANCE 5 [2014]](https://substackcdn.com/image/fetch/w_1300,h_650,c_fill,f_webp,q_auto:good,fl_progressive:steep,g_auto/https%3A%2F%2Fsubstack-video.s3.amazonaws.com%2Fvideo_upload%2Fpost%2F154701811%2F337cccef-f16a-42bf-a6ec-2698fda1b2bb%2Ftranscoded-1739278333.png)
Το νερό έχει μνήμη; Μήπως το στοιχείο που είναι η προέλευση της ζωής έχει επίσης δική του ζωή;
Στο τελευταίο επεισόδιο της 21ης σειράς, η ομάδα του Voyager ερευνά πώς λειτουργεί το DNA μας και πώς τα γονίδιά μας μπορούν να μεταδίδουν πληροφορίες ακριβώς μέσω του νερού. Στο Παρίσι, ο Ρομπέρτο Τζιάκομπο συναντά αποκλειστικά τον νομπελίστα για την ιατρική Λυκ Μοντανιέ, έναν σπουδαίο επιστήμονα που διεξάγει μια μοναδική έρευνα, η οποία θα μπορούσε να ανατρέψει πολλές από τις αρχές της σύγχρονης χημείας και βιολογίας.
Η υπόθεση του Μοντανιέ, στην πραγματικότητα, είναι ότι το DNA μας μπορεί να μεταδοθεί σε μεγάλη απόσταση μέσω ηλεκτρομαγνητικών κυμάτων που διαδίδονται μέσω του νερού. Μια ιδέα που προκάλεσε πραγματικό σεισμό στην επιστημονική κοινότητα, ακριβώς λόγω του κύρους του προσώπου που την πρότεινε.
Το Voyager συγκέντρωσε και συνέκρινε τις απόψεις άλλων επιστημόνων, συμπεριλαμβανομένων δύο επιφανών Ιταλών φυσικών, για να κατανοήσει τις βασικές αρχές πίσω από αυτή την έρευνα και ποιες θα ήταν οι συνέπειες, σε πρακτικό και θεωρητικό επίπεδο, μιας τέτοιας ανακάλυψης.
Κβαντική τηλεμεταφορά του DNA
Μία από τις τελευταίες καινοτομίες στον τομέα της κβαντικής φυσικής είναι το πείραμα που πραγματοποιήθηκε από τον Λυκ Μοντανιέ, βραβευμένο με Νόμπελ ιατρικής το 2008, ο οποίος ανακάλυψε την ικανότητα του DNA να στέλνει ένα είδος ηλεκτρομαγνητικού αποτυπώματος του εαυτού του σε υγρά και κύτταρα.

Ο Jeff Reimers, από το Πανεπιστήμιο του Σίδνεϊ, δηλώνει ότι εάν τα αποτελέσματα αυτού του πειράματος ήταν συγκεκριμένα, αυτή η ανακάλυψη θα έφερνε επανάσταση στις έννοιες της σύγχρονης χημείας, κάνοντας τον δρόμο της ανάμεσα στα πιο σημαντικά πειράματα των τελευταίων 90 ετών.
Πολλοί ερευνητές έχουν αντιδράσει με σκεπτικισμό στην είδηση αυτού του πειράματος, υποστηρίζοντας ότι δεν υπάρχουν διαθέσιμα στοιχεία που να αποδεικνύουν την αλήθεια αυτής της ανακάλυψης.
Τα μόνα διαθέσιμα δεδομένα προς το παρόν είναι η διεξαγωγή αυτής της διαδικασίας στο εργαστήριο, η οποία πραγματοποιείται με τη χρήση δύο παρακείμενων αλλά φυσικά χωριστών σωλήνων που έχουν τοποθετηθεί μέσα σε ένα χάλκινο πηνίο και υπόκεινται σε πολύ ασθενή ηλεκτρομαγνητική συχνότητα. Η συσκευή έχει απομονωθεί από το επίγειο ηλεκτρομαγνητικό πεδίο για την αποφυγή παρεμβολών. Ο πρώτος σωλήνας περιείχε ένα θραύσμα DNA μήκους περίπου 100 βάσεων. το δεύτερο περιείχε καθαρό νερό.
Μετά από 16-18 ώρες, και τα δύο δείγματα υποβλήθηκαν ανεξάρτητα σε μια αλυσιδωτή αντίδραση πολυμεράσης (PCR), μια μέθοδο που χρησιμοποιείται συνήθως για την ενίσχυση ιχνών DNA χρησιμοποιώντας ένζυμα (DNA-πολυμεράση) για τη δημιουργία πολλών αντιγράφων του αρχικού γενετικού υλικού. Το γενετικό θραύσμα ανιχνεύθηκε και στους δύο σωλήνες, παρόλο που ο δεύτερος περιείχε μόνο νερό.
Αυτό το φαινόμενο εμφανίζεται μόνο όταν το αρχικό διάλυμα DNA είχε υποβληθεί σε αρκετούς γύρους αραίωσης πριν τοποθετηθεί μέσα στο μαγνητικό πεδίο. Σε κάθε δοκιμή, το DNA αραιώνεται σε αναλογία 1 προς 10, και αυτό που ονομάστηκε "φανταστικό DNA" ανακτήθηκε μόνο μετά από 7-12 αραιώσεις του αρχικού, και όχι σε ακραίες συγκεντρώσεις παρόμοιες με αυτές που χρησιμοποιούνται για την "ομοιοπαθητική".
Ο Felix Franks, ένας χημικός που έχει μελετήσει το νερό κατά τη διάρκεια της ακαδημαϊκής του σταδιοδρομίας, δηλώνει ότι με βάση τις μελέτες του ότι οι αντιδράσεις που προέκυψαν στο πείραμα δεν είναι αποδεκτές καθώς η δομή θα καταστρεφόταν αμέσως, καθώς το νερό δεν έχει μνήμη, επομένως δεν μπορεί να γίνει ένα αποτύπωμα και να το ανακτήσετε αργότερα.
Ένας συνάδελφος του Montagnier Giuseppe Vitiello, του Πανεπιστημίου του Σαλέρνο, υποστηρίζει ότι τα αποτελέσματα είναι αξιόπιστα και ότι είναι πολύ σημαντικό οι άλλες ομάδες να επαναλάβουν το πείραμα.
Ο Λυκ Μοντανιέ ανακοίνωσε ότι στις αρχές Ιανουαρίου 2011, σε ηλικία 78 ετών, θα αναλάβει τη διεύθυνση ενός νέου ερευνητικού ινστιτούτου στο Πανεπιστήμιο Jiaotong στη Σαγκάη. Σχεδιάζει να μελετήσει τα ηλεκτρομαγνητικά κύματα που προέρχονται από εξαιρετικά αραιωμένο DNA διαφόρων παθογόνων, που μπορεί να αποκαλύψει τη βακτηριακή ή ιογενή προέλευση πολλών καταστάσεων, συμπεριλαμβανομένου του αυτισμού και της νόσου του Αλτσχάιμερ.
Δημοσίευση - Κύματα DNA και νερό [DNA waves and water]
L. Montagnier 1,2, J. Aissa2, E. Del Giudice3, C. Lavallee2, A. Tedeschi4, και G. Vitiello5
1 Παγκόσμιο Ίδρυμα για την έρευνα και την πρόληψη του AIDS (UNESCO), Παρίσι, Γαλλία
2 Nanetics Biotecnologies, S.A. 98 rue Albert Calmette, F78350 Jouy-en-Josas, Γαλλία
3 IIB, Διεθνές Ινστιτούτο Βιοφωτονικής, Neuss, Γερμανία
4 WHITE HB, Μιλάνο, Ιταλία
5 Τμήμα Μαθηματικών και Πληροφορικής, Πανεπιστήμιο του Σαλέρνο και INFN, Gruppo Collegato Salerno, I-84100 Salerno, Ιταλία - E-mail: vitiello@sa.infn.it
Περίληψη:
Ορισμένες βακτηριακές και ιικές αλληλουχίες DNA έχουν βρεθεί ότι προκαλούν ηλεκτρομαγνητικά κύματα χαμηλής συχνότητας σε υψηλές υδατικές αραιώσεις. Αυτό το φαινόμενο φαίνεται να προκαλείται από το ηλεκτρομαγνητικό υπόβαθρο του περιβάλλοντος πολύ χαμηλής συχνότητας. Συζητάμε αυτό το φαινόμενο στο πλαίσιο της κβαντικής θεωρίας πεδίου [quantum field theory]. Προτείνεται ένα σχήμα ικανό να λαμβάνει υπόψη τις παρατηρήσεις. Το αναφερόμενο φαινόμενο θα μπορούσε να επιτρέψει την ανάπτυξη εξαιρετικά ευαίσθητων συστημάτων ανίχνευσης για χρόνιες βακτηριακές και ιογενείς λοιμώξεις.
1. Εισαγωγή: Τα τελευταία 60 χρόνια, η ανάπτυξη βασικών γνώσεων στη βιολογία καθώς και πολλές ιατρικές εφαρμογές οφείλονται κατά πολύ στις ανακαλύψεις που έγιναν στο DNA. Ακολουθεί μια μερική λίστα που δίνει έμφαση στις κύριες προόδους στην ανακάλυψη DNA:
1944 Μετασχηματισμός βακτηρίων από DNA (O. Avery, C. McLeod και M. McCarty)
1953 Αποσαφηνίζεται η δομή της διπλής έλικας (J. Watson, F. Crick, M. Wilkins, R. Franklin) 1956 DNA πολυμεράση (A. Kornberg)
1968 Περιοριστικά ένζυμα (W. Arber)
1969 Αντίστροφη μεταγραφή ρετροϊών (H. Temin, D. Baltimore)
1976 Προσδιορισμός αλληλουχίας DNA (A. Maxam, W. Gilbert, F. Sanger)
1986-1988 Αλυσιδωτή αντίδραση πολυμεράσης (K. Mullis), Taq polymerase (RK Saiki)
2001 Πρώτη αλληλουχία ανθρώπινου γονιδιώματος
2004-2010 Υψηλής απόδοσης αλληλούχιση DNA [High-Throughput DNA Sequencing].
Από την άλλη πλευρά, την ίδια εποχή έχουν συσσωρευτεί στοιχεία για την επίδραση των ηλεκτρομαγνητικών πεδίων (em) στους ζωντανούς οργανισμούς. Οι συχνότητες των εμπλεκόμενων πεδίων em καλύπτουν διαφορετικά διαστήματα (φάσματα) που αντιστοιχούν στις διαφορετικές κλίμακες (διαστάσεις) που υπάρχουν στους οργανισμούς.
Στην παρούσα εργασία, με αναφορά σε πρόσφατα δημοσιευμένα πειραματικά αποτελέσματα [1, 2, 3], συζητάμε την εμφάνιση μιας νέας ιδιότητας του DNA που σχετίζεται με την επαγωγή [induction] πεδίων em εξαιρετικά χαμηλής συχνότητας (ELF). Αυτά τα πεδία μπορούν να προκληθούν με κατάλληλες διαδικασίες σε αραιώσεις νερού που καθίστανται ικανές να διαδώσουν τις πληροφορίες που περιέχονται στο DNA των αρχικών οργανισμών σε άλλους.
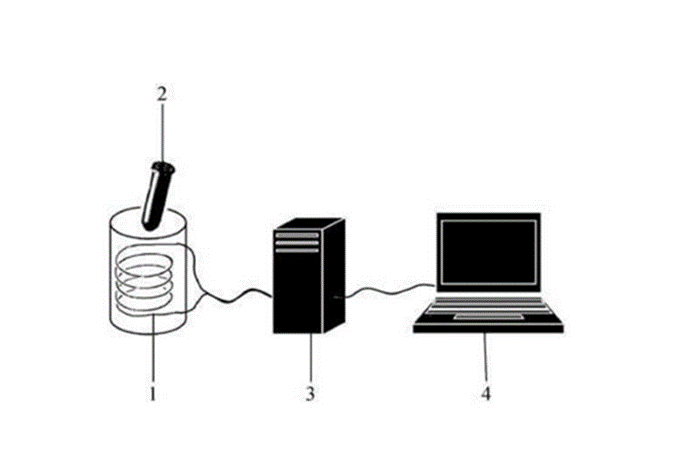
Εικόνα 1. Συσκευή λήψης και ανάλυσης σημάτων em. (1) Πηνίο κατασκευασμένο από σύρμα χαλκού, σύνθετη αντίσταση 300 Ohms. (2) Πλαστικός σωλήνας με πώμα που περιέχει 1 ml του προς ανάλυση διαλύματος. (3) Ενισχυτής. (4) Υπολογιστής. Από την Αναφ. 1
Η εργασία περιλαμβάνει τρεις ενότητες: τα νέα δεδομένα, ένα θεωρητικό σχήμα όπου θα τα συζητήσουμε και τις ιατρικές εφαρμογές.
2. Τα νέα δεδομένα: μια νέα ιδιότητα του DNA και η επαγωγή ηλεκτρομαγνητικών κυμάτων σε αραιώσεις νερού.
Η ιστορία ξεκίνησε πριν από δέκα χρόνια όταν ένας από εμάς (LM [Luc Montagnier]) μελέτησε την παράξενη συμπεριφορά ενός μικρού βακτηρίου, συχνού συντρόφου του HIV, το Mycoplasma pirum, και όπως ο HIV είναι λάτρης των ανθρώπινων λεμφοκυττάρων. Ο LM προσπαθούσε να διαχωρίσει το βακτήριο, το οποίο έχει μέγεθος περίπου 300 nm, από τα ιικά σωματίδια των οποίων το μέγεθος είναι περίπου 120 nm με διήθηση χρησιμοποιώντας φίλτρα των 100 nm και 20 nm.
Ξεκινώντας με καθαρές καλλιέργειες του βακτηρίου σε λεμφοκύτταρα, τα διηθήματα ήταν πράγματι στείρα από το βακτήριο όταν καλλιεργήθηκαν σε ένα πλούσιο κυτταρικό μέσο, SP4. Η αλυσιδωτή αντίδραση πολυμεράσης (PCR) και η ένθετη PCR, με βάση εκκινητές προερχόμενους από ένα γονίδιο του M. pirum που είχε προηγουμένως κλωνοποιηθεί και προσδιοριστεί η αλληλουχία, η συγκολλητίνη, ήταν αρνητική στο διήθημα. Ωστόσο, όταν το διήθημα καλλιεργήθηκε με ανθρώπινα λεμφοκύτταρα, (με προηγούμενο έλεγχο ότι δεν είχε μολυνθεί από το μυκόπλασμα) το μυκόπλασμα με όλα τα χαρακτηριστικά του επανήλθε κανονικά!
Τότε τέθηκε το ερώτημα: τι είδους πληροφορίες μεταδόθηκαν στο υδατικό διήθημα; Ήταν η αρχή μιας μακροχρόνιας έρευνας που αφορούσε τις φυσικές ιδιότητες του DNA στο νερό. Πράγματι, βρέθηκε μια νέα ιδιότητα του DNA του M. pirum: η εκπομπή κυμάτων χαμηλής συχνότητας σε ορισμένες αραιώσεις νερού του διηθήματος, και σύντομα επεκτάθηκε και σε άλλα βακτηριακά και ιικά DNA.
Η συσκευή που χρησιμοποιείται για την ανίχνευση των ηλεκτρομαγνητικών σημάτων περιλαμβάνει ένα σωληνοειδές (πηνίο) που λαμβάνει τη μαγνητική συνιστώσα των κυμάτων που παράγονται από το διάλυμα DNA σε έναν πλαστικό σωλήνα που μετατρέπει τα σήματα σε ηλεκτρικό ρεύμα. Αυτό το ρεύμα στη συνέχεια ενισχύεται και τελικά αναλύεται σε φορητό υπολογιστή χρησιμοποιώντας συγκεκριμένο λογισμικό (Εικ. 1).
Ακολουθεί μια σύντομη περίληψη των εργαστηριακών παρατηρήσεων, οι οποίες περιγράφονται λεπτομερέστερα στο [1, 2, 3]:
1) Ηλεκτρομαγνητικά κύματα εξαιρετικά χαμηλής συχνότητας (ULF 500 – 3.000 Hz) ανιχνεύθηκαν σε ορισμένες αραιώσεις διηθημάτων (100 nm, 20 nm) από καλλιέργειες μικροοργανισμών (ιός, βακτήρια) ή από το πλάσμα ανθρώπων που έχουν μολυνθεί με τους ίδιους παράγοντες (Εικ. 2). Τα ίδια αποτελέσματα λαμβάνονται από το εξαγόμενο DNA τους.
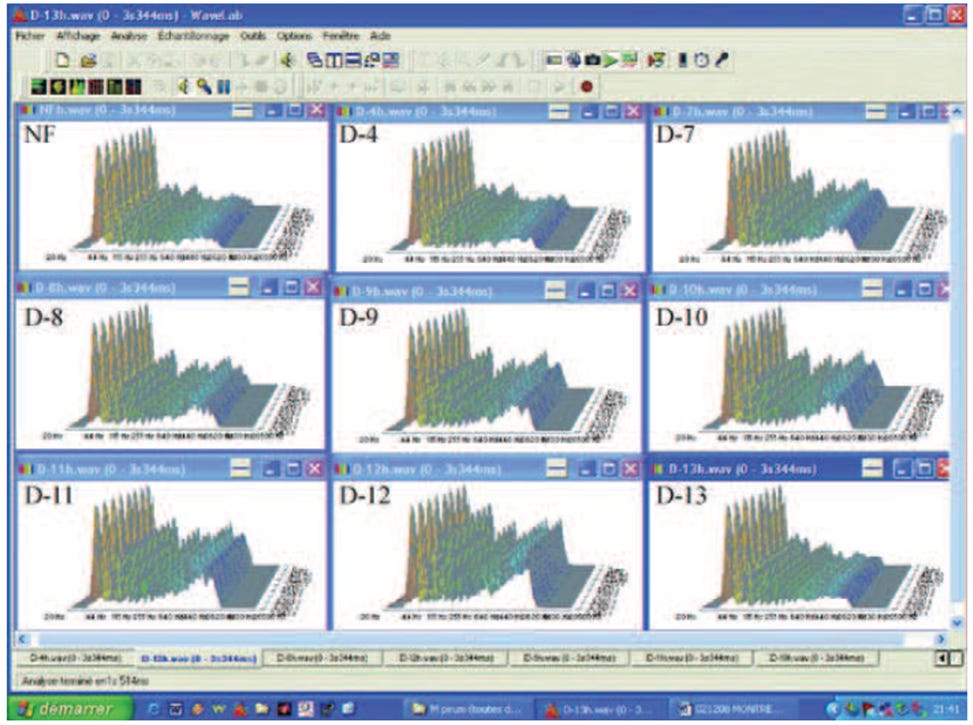
Σχήμα 2. Τυπικά σήματα από υδατικές αραιώσεις του M. pirum (λογισμικό Matlab). Σημειώστε τα θετικά σήματα από τις αραιώσεις D-7 έως D-12. Από το σημείο αναφοράς 1.
2) Τα ηλεκτρομαγνητικά σήματα (EMS) δεν συσχετίζονται γραμμικά με τον αρχικό αριθμό των βακτηριακών κυττάρων πριν από τη διήθησή τους. Σε ένα πείραμα τα EMS ήταν παρόμοια σε ένα εναιώρημα κυττάρων E. coli που κυμαίνονταν από 109 έως 10. Είναι ένα φαινόμενο όλα ή κανένα.
3) EMS παρατηρούνται μόνο σε ορισμένες υψηλές αραιώσεις νερού των διηθημάτων. Για παράδειγμα, από 10^9 έως 10^18 αραιώσεις σε ορισμένα παρασκευάσματα διηθημάτων E. coli.
4) Στην περίπτωση του M. pirum, ένα απομονωμένο, μεμονωμένο γονίδιο (συγκολλητίνη, προηγουμένως κλωνοποιημένη και προσδιορισμένη αλληλουχία) θα μπορούσε να προκαλέσει το EMS. Καθώς το γονίδιο κλωνοποιήθηκε σε δύο θραύσματα, καθένα από τα απομονωμένα θραύσματα ήταν σε θέση να δημιουργήσει EMS, υποδηλώνοντας ότι μια σύντομη αλληλουχία DNA ήταν αρκετή για να επάγει τα σήματα. Παρομοίως, μια σύντομη αλληλουχία DNA HIV (104 ζεύγη βάσεων) βρέθηκε ότι είναι επαρκής για την παραγωγή του EMS.
5) Ορισμένα βακτήρια δεν παράγουν EMS: αυτή είναι η περίπτωση προβιοτικών βακτηρίων όπως π.χ. το Lactobacillus και επίσης ορισμένων εργαστηριακών στελεχών E. coli που χρησιμοποιούνται ως φορέας κλωνοποίησης.
6) Αυτές οι μελέτες έχουν επεκταθεί στους ιούς, αν και δεν έχουν διερευνηθεί όλες οι οικογένειες ιών. Παρόμοια EMS ανιχνεύθηκαν από ορισμένους εξωγενείς ρετροϊούς (HIV, FeLV), ιούς ηπατίτιδας (HBV, HCV) και γρίπης Α (καλλιέργειες in vitro). Γενικά, τα EMS παράγονται από διηθήματα 20 nm ιϊκών εναιωρημάτων ή από το εξαγόμενο DNA: παραμένει ένα ερώτημα για τους ιούς RNA (HCV, γρίπη) ως προς το εάν το RNA από τα ώριμα ιικά σωματίδια είναι πηγή EMS ή όχι.
Στην περίπτωση του HIV, τα EMS δεν παράγονται από το RNA των ιϊκών σωματιδίων, αλλά μάλλον παράγονται από το προϊικό DNA που υπάρχει στα μολυσμένα κύτταρα. Στην περίπτωση των βακτηρίων, το EMS παράγεται από διηθήματα 100 nm και όχι από διηθήματα 20 nm, υποδεικνύοντας ότι το μέγεθος των δομών που παράγουν EMS κυμαίνεται μεταξύ 20 και 100 nm.
Αυτό δικαιολογεί το όνομα των νανοδομών. Αυτές οι μελέτες υποδηλώνουν ότι έχουμε να κάνουμε με νανοδομές από νερό. Χρησιμοποιούνται δείγματα νερού υψηλής καθαρότητας, αν και δεν μπορεί να αποκλειστεί ο ρόλος ελάχιστων ιχνών ακαθαρσιών.
Η παραγωγή EMS από τις νανοδομές είναι ανθεκτική σε: επεξεργασία RNAse, DNase (ενώ αυτό θα καταστρέψει το DNA από την έναρξη του EMS), Πρωτεάση (πρωτεϊνάση K), Αποστειρωτικό (SDS). Ωστόσο, είναι ευαίσθητα στη θερμότητα (πάνω από 70 C) και στον παγετό ( 80 C).
Αυτή η ευαισθησία μειώνεται όταν έχουμε να κάνουμε με καθαρισμένες βραχείες αλληλουχίες DNA.
Οι τεχνικές συνθήκες για την επαγωγή του EMS συνοψίζονται στην ακόλουθη λίστα:
- Φιλτράρισμα 450/100 nm για βακτηριακό DNA, 450/20 nm για ιικό DNA
- Υψηλές αραιώσεις σε νερό
- Μηχανική ανάδευση (Vortex) μεταξύ κάθε αραίωσης
- Διέγερση με ηλεκτρομαγνητικό υπόβαθρο εξαιρετικά χαμηλής συχνότητας (ELF), ξεκινώντας από πολύ χαμηλά στα 7 Hz. Η διέγερση δεν προκαλείται όταν το σύστημα θωρακίζεται από ένα κλωβό από μ-μέταλλο.
- Η διέγερση από το ηλεκτρομαγνητικό υπόβαθρο πολύ χαμηλής συχνότητας είναι απαραίτητη. Το (ηλεκτρομαγνητικό) υπόβαθρο παράγεται είτε από φυσικές πηγές (οι συντονισμοί Schumann [4] που ξεκινούν στα 7,83 Hz) είτε από τεχνητές πηγές.
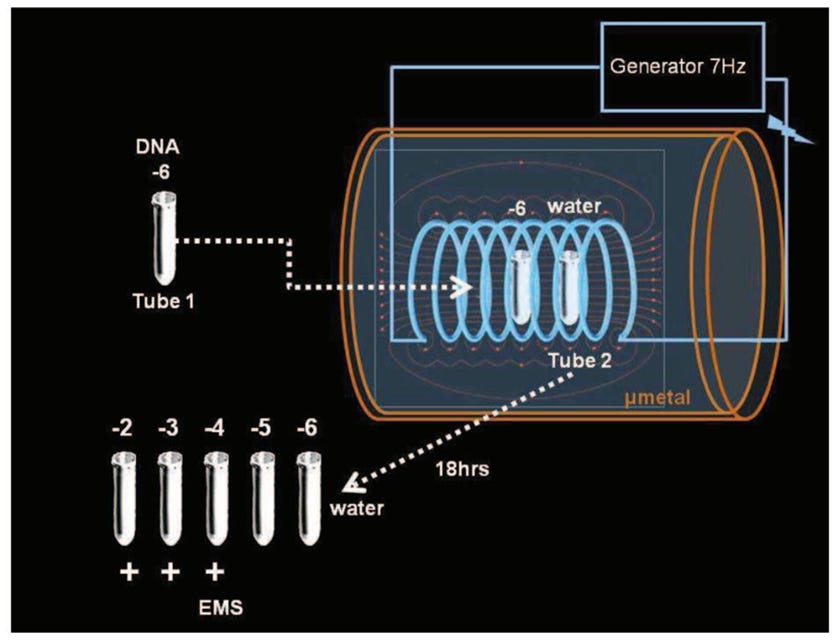
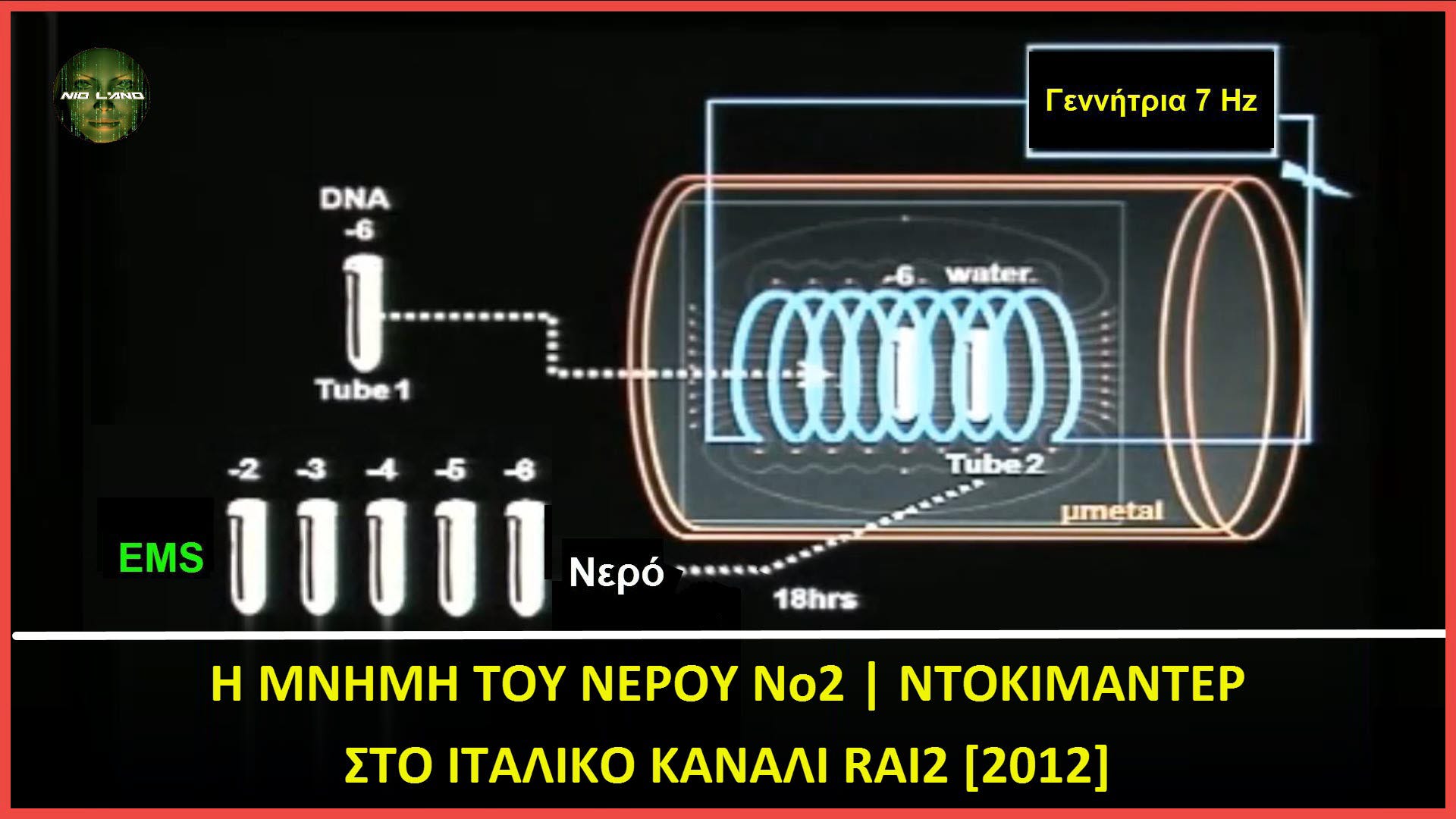
Εικόνα 3. Μετάδοση γενετικής πληροφορίας DNA στο νερό μέσω ηλεκτρομαγνητικών κυμάτων. Από την Αναφ. 3
2.1. Μετάδοση της αλληλουχίας DNA μέσω κυμάτων και νερού
Σε περαιτέρω πειράματα ένα θραύσμα DNA του HIV που ελήφθη από τη μακρά τελική επανάληψη του (LTR - long terminal repeat) έχει χρησιμοποιηθεί ως πηγή DNA. Αυτό το θραύσμα ενισχύθηκε με PCR (487 ζεύγη βάσεων) και ένθετη PCR (104 ζεύγη βάσεων) χρησιμοποιώντας ειδικούς εκκινητές.
Σε ένα πρώτο βήμα, έγιναν αραιώσεις DNA, στις οποίες ανιχνεύθηκε η παραγωγή EMS κάτω από το ηλεκτρομαγνητικό υπόβαθρο του περιβάλλοντος.
Στη συνέχεια έγιναν τα ακόλουθα βήματα. Όπως φαίνεται στο Σχ. 3, μία από τις θετικές αραιώσεις (ας πούμε 10^(-6) ) τοποθετήθηκε σε ένα κουτί προστατευόμενο από στρώμα μ-μετάλλου πάχους 1 mm (ένα κράμα που απορροφά κύματα εξαιρετικά χαμηλής συχνότητας). Κοντά του τοποθετήθηκε ένας άλλος σωλήνας που περιείχε καθαρό νερό. Η περιεκτικότητα σε νερό κάθε σωλήνα διηθήθηκε μέσω φίλτρων 450 nm και 20 nm και αραιώθηκε από 10^(-2) έως 10^(-15). Γύρω τους τοποθετείται ένα πηνίο χαλκού που δέχεται ηλεκτρικό ρεύμα χαμηλής έντασης και ταλάντωσης στα 7 Hz, που παράγεται από μια εξωτερική γεννήτρια. Το παραγόμενο μαγνητικό πεδίο διατηρείται για 18 ώρες σε θερμοκρασία δωματίου. Στη συνέχεια, το EMS καταγράφεται από κάθε σωλήνα. Τώρα και ο σωλήνας που περιέχει νερό εκπέμπει EMS, στις αραιώσεις που αντιστοιχούν σε εκείνες που είναι θετικές για EMS στον αρχικό σωλήνα DNA.
Αυτό το αποτέλεσμα δείχνει ότι, με διέγερση 7 Hz, έχει επιτευχθεί η μετάδοση σε καθαρό νερό της ταλάντωσης των νανοδομών που αρχικά προήλθαν από το DNA.
Οι ακόλουθοι έλεγχοι βρέθηκαν να καταστέλλουν τη μετάδοση του EMS στο σωλήνα νερού:
- Χρόνος έκθεσης των δύο σωλήνων μικρότερος από 16 - 18 ώρες
- Χωρίς πηνίο
- Απενεργοποιημένη γεννήτρια μαγνητικού πεδίου
- Συχνότητα διέγερσης < 7 Hz
- Απουσία DNA στον σωλήνα 1.
Σε αυτό το σημείο έγινε το πιο κρίσιμο βήμα, δηλαδή η διερεύνηση της ιδιομορφίας
των επαγόμενων νανοδομών νερού, με την αναδημιουργία από αυτές της αλληλουχίας DNA.
Για αυτό όλα τα συστατικά για τη σύνθεση του DNA με αλυσιδωτή αντίδραση πολυμεράσης (νουκλεοτίδια, εκκινητές, πολυμεράση) προστέθηκαν στον σωλήνα του νερού (που δέχτηκε) με το σήμα.
Η ενίσχυση πραγματοποιήθηκε υπό κλασικές συνθήκες (35 κύκλοι) σε θερμοκυκλοποιητή. Το DNA που παρήχθη στη συνέχεια υποβλήθηκε σε ηλεκτροφόρηση σε γέλη (gel) αγαρόζης. Το αποτέλεσμα ήταν ότι ανιχνεύθηκε μια ζώνη DNA του αναμενόμενου μεγέθους του αρχικού θραύσματος LTR. Επιβεβαιώθηκε περαιτέρω ότι αυτό το DNA είχε μια αλληλουχία πανομοιότυπη ή σχεδόν ίδια με την αρχική αλληλουχία DNA του LTR. Στην πραγματικότητα, ήταν 98 % πανομοιότυπο (διαφορά 2 νουκλεοτιδίων) από τα 104. Αυτό το πείραμα βρέθηκε να είναι εξαιρετικά αναπαραγώγιμο (12 από 12) και επαναλήφθηκε επίσης με μια άλλη αλληλουχία DNA από ένα βακτήριο, το Borrelia burgdorferi, τον παράγοντα της νόσου του Lyme.
Φάνηκε ξεκάθαρα ότι οι νανοδομές του νερού και ο ηλεκτρομαγνητικός συντονισμός τους μπορούν να διαιωνίσουν (αναπαραγάγουν) πιστά τις πληροφορίες DNA.
Αυτά τα στοιχεία υποστηρίζουν μια προκλητική εξήγηση του πειράματος διήθησης Mycoplasma pirum (Εικ. 1): οι νανοδομές που προκαλούνται από το DNA του M. pirum στο φιλτραρισμένο νερό αντιπροσωπεύουν διαφορετικά τμήματα του γονιδιωματικού DNA του. Κάθε νανοδομή όταν έρχεται σε επαφή με τα ανθρώπινα λεμφοκύτταρα μεταγράφεται εκ των υστέρων στο αντίστοιχο DNA από ορισμένες κυτταρικές πολυμεράσες DNA.
Τα στοιχεία αυτά υποστηρίζουν μια ανατρεπτική εξήγηση του πειράματος διήθησης του Mycoplasma pirum (Εικ. 1): οι νανοδομές που προκαλούνται από το DNA του M. pirum στο διηθημένο (φιλτραρισμένο) νερό αντιπροσωπεύουν διαφορετικά τμήματα του γονιδιωματικού DNA του. Κάθε νανοδομή όταν έρχεται σε επαφή με τα ανθρώπινα λεμφοκύτταρα ανα-μεταγράφεται (μεταγράφεται εκ των υστέρων) στο αντίστοιχο DNA από κάποιες πολυμεράσες του κυτταρικού DNA.
Στη συνέχεια, υπάρχει μια ορισμένη πιθανότητα (έστω και πολύ μικρή) να ανασυνδυαστεί κάθε κομμάτι DNA μέσα στο ίδιο κύτταρο με άλλα κομμάτια για την ανακατασκευή ολόκληρου του γονιδιώματος του DNA. Πρέπει να υποθέσουμε ότι παρουσία ευκαρυωτικών κυττάρων η σύνθεση των συστατικών του μυκοπλάσματος (λιπίδια της μεμβράνης, ριβοσώματα) μπορεί επίσης να καθοδηγείται από το DNA του μυκοπλάσματος. Ένα μόνο πλήρες μυκοπλασματικό κύτταρο είναι τότε αρκετό για να δημιουργήσει ολόκληρη τη μόλυνση των λεμφοκυττάρων.
Πρόσφατα πειράματα της ομάδας G. Vinter έδειξαν [5] ότι ένα συνθετικό γονιδιωματικό DNA επαρκεί για να διατηρήσει όλα τα χαρακτηριστικά ενός μυκοπλάσματος. Όλα τα στάδια που απαιτούνται για την ανασύνθεση από το νερό μπορούν να αναλυθούν και να επαληθευτούν.
3. Το θεωρητικό πλαίσιο
Οι παραπάνω πειραματικές παρατηρήσεις ταιριάζουν στη φυσική άποψη που αντιμετωπίζει τη βιολογική δυναμική ως αλληλεπίδραση χημικών διεργασιών και αλληλεπιδράσεων. με άλλα λόγια, ως μια σειρά από υποβοηθούμενες βιοχημικές αντιδράσεις.
Θα προσπαθήσουμε να ερμηνεύσουμε τα παραπάνω πειραματικά αποτελέσματα στο πλαίσιο μιας πρόσφατα προτεινόμενης θεωρίας για το υγρό νερό που βασίζεται στην Κβαντική Θεωρία Πεδίου (QFT - Quantum Field Theory) [6]-[11]. Αυτή η θεωρία είναι εγγενώς μη γραμμική και παρέχει τα κατάλληλα εργαλεία για να περιγράψει ένα πολύπλοκο σύνολο διαδικασιών που είναι επίσης μη γραμμικές.
Ας συνοψίσουμε πρώτα τα κύρια σημεία μιας τέτοιας θεωρίας, της οποίας οι λεπτομέρειες μπορούν να βρεθούν στις αναφερόμενες παραπομπές.
Το σημείο εκκίνησης είναι η συνειδητοποίηση ότι τα μόρια του υγρού νερού δεν μπορεί να θεωρηθεί ότι συνδέονται με αμιγώς στατικές αλληλεπιδράσεις (δεσμοί Η, ηλεκτρική αλληλεπίδραση διπόλου-διπόλου). Η σύνδεσή τους προκαλείται στην πραγματικότητα από το εξαρτώμενο από το χρόνο ακτινοβολούμενο πεδίο em μακρινού πεδίου. Οι στατικοί δεσμοί κοντινού πεδίου, όπως οι δεσμοί Η, δημιουργούνται στη συνέχεια ως συνέπεια της συμπύκνωσης των μορίων που προκαλείται από τα εν λόγω πεδία ακτινοβολίας μακρινού πεδίου.
Τα κυριότερα αποτελέσματα της θεωρίας είναι [6]-[11]:
α) Ένα σύνολο μορίων που αλληλεπιδρούν με το πεδίο ακτινοβολίας em αποκτά, πάνω από ένα κατώφλι πυκνότητας και κάτω από μια κρίσιμη θερμοκρασία, μια νέα μη τετριμμένη κατάσταση ελάχιστης ενέργειας, διαφορετική από τη συνήθη κατάσταση όπου οι ταλαντώσεις των μορίων είναι ασυσχέτιστες (ασύνδετες) και το πεδίο em εξαφανίζεται. Η νέα κατάσταση ελάχιστης ενέργειας συνεπάγεται μια διαμόρφωση του συστήματος όπου όλα τα μόρια που περικλείονται σε μια εκτεταμένη περιοχή, η οποία ονομάζεται περιοχή συνοχής (Coherence Domain - CD), ταλαντώνονται σε αρμονία με ένα πεδίο em ηλεκτρομαγνητικής ακτινοβολίας που είναι παγιδευμένο στο εσωτερικό του CD.
Το μέγεθος αυτής της εκτεταμένης περιοχής είναι μόνο το μήκος κύματος λ του παγιδευμένου πεδίου em. Η συλλογική συνεκτική ταλάντωση του συστατικού των μορίων του CD λαμβάνει χώρα μεταξύ της βασικής κατάστασης του μεμονωμένου μορίου και μιας διεγερμένης κατάστασης της οποίας ο όγκος, σύμφωνα με την ατομική φυσική, είναι ευρύτερος από τον όγκο θεμελιώδους κατάστασης. Το μήκος κύματος του παγιδευμένου πεδίου em εξαρτάται από την ενέργεια διέγερσης Eexc μέσω την εξίσωσης [λ = hc / Eexc]:
Το μέγεθος αυτής της διευρυμένης περιοχής είναι ακριβώς το μήκος κύματος λ του εγκλωβισμένου (παγιδευμένου) em πεδίου. Η συλλογική συνεκτική ταλάντωση των μορίων που συνιστούν το CD λαμβάνει χώρα μεταξύ της βασικής κατάστασης του μεμονωμένου μορίου και μιας διεγερμένης κατάστασης της οποίας ο όγκος, σύμφωνα με την ατομική φυσική, είναι ευρύτερος από τον όγκο της βασικής (θεμελιώδους) κατάστασης. Το μήκος κύματος λ του παγιδευμένου πεδίου em εξαρτάται από την ενέργεια διέγερσης Eexc μέσω της εξίσωσης [λ = hc / Eexc]:

Το CD είναι μια αυτοπαραγόμενη κοιλότητα [cavity] για το πεδίο em λόγω του γνωστού μηχανισμού Anderson-Higgs-Kibble [9], ο οποίος συνεπάγεται ότι το φωτόνιο του παγιδευμένου πεδίου em αποκτά φανταστική μάζα και συνεπώς αδυνατεί να εγκαταλείψει το CD.
Αυτή ακριβώς η αυτοπαγίδευση του em πεδίου εγγυάται ότι η ενέργεια του CD έχει ένα πεπερασμένο κατώτερο όριο. Εξαιτίας αυτής της αυτοπαγίδευσης η συχνότητα του πεδίου em του CD γίνεται πολύ μικρότερη από τη συχνότητα του ελεύθερου πεδίου που έχει το ίδιο μήκος κύματος. Τα παραπάνω αποτελέσματα ισχύουν για όλα τα υγρά.
Η ιδιαιτερότητα του νερού είναι ότι η συνεκτική ταλάντωση λαμβάνει χώρα μεταξύ της βασικής (θεμελιώδους) κατάστασης και μιας διεγερμένης κατάστασης που βρίσκεται στα 12,06 eV ακριβώς κάτω από το κατώφλι ιονισμού (12,60 eV). Στην περίπτωση του υγρού νερού, το CD (του οποίου το μέγεθος είναι 100 nm σύμφωνα με την εξίσωση (1)) περιλαμβάνει ένα σύνολο σχεδόν ελεύθερων ηλεκτρονίων τα οποία είναι σε θέση να δέχονται εξωτερικά παρεχόμενη ενέργεια και να τη μετατρέπουν σε συνεκτικές διεγέρσεις (δίνες) των οποίων η εντροπία είναι πολύ μικρότερη από την εντροπία της εισερχόμενης ενέργειας. Κατά συνέπεια, τα CD του νερού θα μπορούσαν να γίνουν δομές διάχυσης [dissipative structures] κατά την έννοια της θερμοδυναμικής των μη αντιστρεπτών διεργασιών [12]-[14].
β) Η συνοχή μεταξύ των μορίων εξουδετερώνεται σε οποιαδήποτε θερμοκρασία Τ που είναι αμετάβλητη (non-vanishing) με θερμικές συγκρούσεις που θα μπορούσαν να θέσουν τα μόρια εκτός συντονισμού, όπως στην εικόνα Landau του υγρού Ηλίου [15]. Ο ανταγωνισμός μεταξύ της ηλεκτροδυναμικής έλξης και του θερμικού θορύβου παράγει ένα μόνιμο πέρασμα των μορίων μεταξύ ενός συνεκτικού και ενός μη συνεκτικού πλαισίου.
Για μια δεδομένη τιμή του Τ, ο συνολικός αριθμός των συνεκτικών και μη συνεκτικών μορίων είναι σταθερός, αλλά κάθε μόριο ταλαντώνεται μεταξύ των δύο καταστάσεων δημιουργώντας μια συνεχή αλλαγή της κατανομής του χώρου των συνεκτικών και μη συνεκτικών κλασμάτων των μορίων. Αυτό ακριβώς το ταλαντευόμενο περιβάλλον των δύο φάσεων του υγρού νερού είναι που παράγει, στην περίπτωση πειραμάτων των οποίων ο χρόνος ανάλυσης είναι αρκετά μεγάλος, την εμφάνιση του νερού ως ομοιογενούς υγρού. Ωστόσο, η παραπάνω ιδιότητα ισχύει μόνο για το ελεύθερο νερό.
Κοντά σε μια επιφάνεια, η έλξη μεταξύ των μορίων του νερού και της επιφάνειας θα μπορούσε να προστατεύσει τη συνεκτική δομή από τον θερμικό θόρυβο, προκαλώντας μια σταθεροποίηση της συνεκτικής δομής.
Αυτό ισχύει ιδίως στην περίπτωση των ζωντανών οργανισμών, όπου τα μόρια του νερού οριοθετούνται με μεμβράνες ή σκελετούς βιομορίων. Σε αυτή την περίπτωση τα CDs επιβιώνουν αρκετά ώστε να παρουσιάζουν τις ιδιαίτερες ιδιότητες της συνοχής.
γ) Τα CD αποθηκεύουν την εξωτερικά παρεχόμενη ενέργεια με τη μορφή συνεκτικών στροβιλισμών. Αυτές οι δίνες έχουν μακρά διάρκεια λόγω της συνοχής, έτσι ώστε μια μόνιμη εισροή ενέργειας να παράγει μια συσσώρευση στροβιλισμών- αθροίζονται για να δώσουν μια μοναδική δίνη της οποίας η ενέργεια είναι το άθροισμα των μερικών ενεργειών των διεγέρσεων που έχουν αθροιστεί. Με αυτόν τον τρόπο τα CD του νερού μπορούν να αποθηκεύσουν ένα σημαντικό ποσό ενέργειας σε μια μοναδική συνεκτική διέγερση ικανή να ενεργοποιήσει τους βαθμούς ελευθερίας των ηλεκτρονίων των μορίων- αυτή η υψηλής ποιότητας ενέργεια είναι το άθροισμα πολλών μικρών συνεισφορών, των οποίων η αρχική εντροπία ήταν υψηλή.
δ) Τα CD ταλαντώνονται σε μια συχνότητα κοινή για το em πεδίο και τα μόρια του νερού και η συχνότητα αυτή αλλάζει όταν αποθηκεύεται ενέργεια στο CD.
Όταν η συχνότητα ταλάντωσης του CD ταιριάζει με τη συχνότητα ταλάντωσης κάποιων μη υδατικών μοριακών ειδών που υπάρχουν στα όρια του CD, αυτά τα "φιλοξενούμενα" μόρια γίνονται μέλη του CD και είναι σε θέση να συλλάβουν ολόκληρη την αποθηκευμένη ενέργεια, η οποία γίνεται ενέργεια ενεργοποίησης των φιλοξενούμενων μορίων - κατά συνέπεια, το CD εκφορτίζεται και μπορεί να ξεκινήσει ένας νέος κύκλος ταλάντωσης.
Ο παραπάνω μηχανισμός ταιριάζει με τη διορατικότητα του Albert Szent-Gyorgyi [16], ο οποίος πρότεινε πριν από μισό αιώνα ότι το νερό που περιβάλλει τα βιομόρια πρέπει να είναι η αιτία των διεγέρσεων των ηλεκτρονίων των μορίων που ευθύνονται για τις χημικές αντιδράσεις.
Επιπλέον, εάν το σύνολο των συχνοτήτων που είναι ικανό να προσελκύσει τα μονομερή συστατικά ενός πολυμερούς διεγερθεί στο CD του νερού, το πολυμερές θα δημιουργηθεί από την έλξη των μονομερών στο CD, υπό την προϋπόθεση ότι αυτά υπάρχουν στο διάλυμα. Με αυτόν τον τρόπο είναι δυνατόν να προκληθεί ο πολυμερισμός των μονομερών με την παροχή στα CD του νερού, του μονομερούς διαλύματος, των em πεδίων που έχουν τις σχετικές συχνότητες (ηλεκτρομαγνητική πληροφορία).
ε) Μια συλλογική απόδοση των CD του νερού, η οποία θα μπορούσε να προκαλέσει μια σύγχρονη βιοχημική δραστηριότητα σε μια μεσοσκοπική περιοχή, θα πρέπει να απαιτεί ομοιόμορφο ρυθμό ενεργειακής φόρτισης για όλα τα εμπλεκόμενα CD. Η απαίτηση αυτή ικανοποιείται από έναν μηχανισμό που περιλαμβάνει ιόντα ηλεκτρολυτών, των οποίων ο ουσιαστικός ρόλος στη βιολογική δυναμική είναι ευρέως αναγνωρισμένος. Τα ιόντα που βρίσκονται κοντά στα CDs του νερού έλκονται από το em πεδίο που παγιδεύεται στις περιοχές - έτσι διατηρούνται σε τροχιά γύρω από την περιοχή κινούμενα με κυκλική ταχύτητα ανάλογη της λεγόμενης συχνότητας c του κυκλοτρονίου:
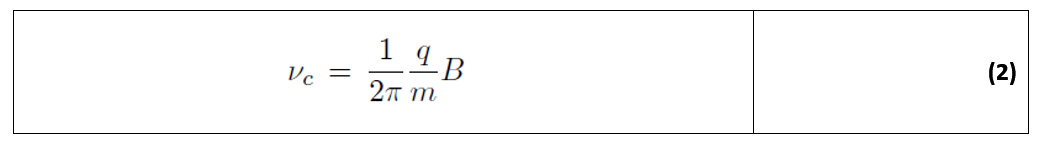
όπου q και m είναι το ηλεκτρικό φορτίο και η μάζα του ιόντος, αντίστοιχα, και Β είναι το μαγνητικό πεδίο.
Δεδομένου ότι το DNA και οι πρωτεΐνες είναι πολυηλεκτρολύτες, περιβάλλονται από ένα σύννεφο θετικών αντίθετων ιόντων, που έχουν συχνότητα κυκλοτρονίου στην περιοχή μεταξύ 1 και 100 Hz και παίζουν σημαντικό ρόλο.
Έχει ανιχνευθεί πειραματικά από τον M. Zhadin [17] και αργότερα από τους Zhadin και Giuliani [18] ότι με την εφαρμογή μαγνητικού πεδίου, με συχνότητα που αντιστοιχεί σε τη συχνότητα του ιοντικού κυκλοτρονίου, σε ένα σύστημα όπου υπάρχουν ιόντα, τα ιόντα αυτά απομακρύνονται από τις τροχιές τους.
Ο μηχανισμός αυτός έχει αποσαφηνιστεί θεωρητικά στο [19]. Λόγω της διατήρησης της στροφορμής (γωνιακής ορμής), η απομάκρυνση των ιόντων από τις τροχιές του κυκλοτρονίου παράγει μια περιστροφική κίνηση των σχεδόν ελεύθερων ηλεκτρονίων των CD του νερού, τα οποία επομένως διεγείρονται ενεργειακά. [20].
Στην περίπτωση που η συγκέντρωση ιόντων θα μπορούσε να θεωρηθεί ομοιόμορφη σε μια μεσοσκοπική περιοχή και το εξωτερικά εφαρμοζόμενο μαγνητικό πεδίο έχει επίσης μεσοσκοπικό μέγεθος, η ποσότητα των ενεργειακής διέγερσης θα μπορούσε να θεωρηθεί ομοιόμορφη σε μια περιοχή που περιλαμβάνει μεγάλο αριθμό CDs νερού τα οποία αντίστοιχα διεγείρονται με ομοιόμορφο τρόπο, εξασφαλίζοντας έτσι τη συνοχή μεταξύ τους.
Η παραμονή στο χρόνο τέτοιων μαγνητικών πεδίων εξαιρετικά χαμηλής συχνότητας εγγυάται μια σταθερή διέγερση των CD νερού και αντίστοιχα της βιοχημικής δραστηριότητας που καταλύεται από αυτά.
Ας αναλύσουμε τώρα τα πειραματικά αποτελέσματα που αναφέρθηκαν στην Ενότητα 2 εντός του θεωρητικού πλαισίου που συνοψίστηκε παραπάνω.
Ο ρόλος που διαδραματίζει το υπόβαθρο της χαμηλής em συχνότητας γίνεται κατανοητός παρατηρώντας ότι για να μεταφερθεί ενέργεια στα CDs του νερού, χρειαζόμαστε ένα εναλλασσόμενο μαγνητικό πεδίο συντονισμού.
Στους ανώτερους οργανισμούς, όπως ο άνθρωπος, το πεδίο αυτό μπορεί να παραχθεί από το νευρικό σύστημα.
Στοιχειώδες οργανισμοί, όπως τα βακτήρια, θα πρέπει να χρησιμοποιούν περιβαλλοντικά πεδία. Καλοί υποψήφιοι είναι το πεδίο Σούμαν [Schumann] του γεωμαγνητικού πεδίου [4].
Αυτοί οι μηχανισμοί είναι οι στατικοί τρόποι (μηχανισμοί) που παράγονται από το μαγνητική δραστηριότητα (αστραπές ή οτιδήποτε άλλο) που λαμβάνει χώρα στη κοιλότητα του οποίου τα όρια είναι η επιφάνεια του Γης και η αγώγιμη ιονόσφαιρα, η οποία λειτουργεί ως κατοπτρικό τοίχωμα για τα μήκη κύματος υψηλότερα από μερικές εκατοντάδες μέτρα.
Αυτοί οι στατικοί τρόποι λειτουργίας θα πρέπει να έχουν συχνότητα νs που στην ιδανική περίπτωση είναι [4]:
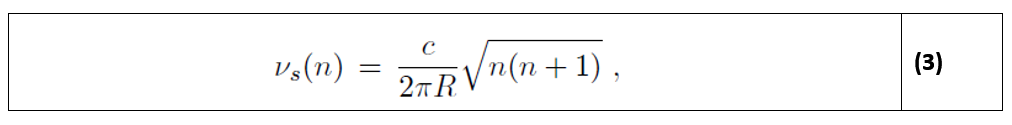
όπου R είναι η ακτίνα της Γης.
Η πραγματική κοιλότητα Γης-ιονόσφαιρας δεν είναι τέλεια, οπότε οι πραγματικές συχνότητες είναι λίγο χαμηλότερες από τις τιμές που δίνει η εξίσωση (3). Στην πραγματικότητα, πειραματικά βρέθηκαν κορυφώσεις γύρω από τα 7,83, 14,3, 20,8, 27,3 και 33,8 Hz.
Κατά συνέπεια, προκειμένου να παραχθεί η ενεργειακή φόρτιση των CDs, το βιολογικό σύστημα θα πρέπει να επιλέγει είδη ιόντων με λόγο q/m τέτοιο ώστε, δεδομένης της τοπικής τιμής του B στον οργανισμό, η τιμή που δίνεται από την Εξ. (2) να ταιριάζει με έναν από τους συντονισμούς Schumann. Η τοπική τιμή του Β αναμένεται να μην διαφέρει πολύ από την τιμή του γήινου μαγνητικού πεδίου, η οποία είναι της τάξης των 50 microtesla.
Όπως εξηγείται στο σημείο ε ανωτέρω, τα ιόντα απομακρύνονται από τις τροχιές τους όταν ένα μαγνητικό πεδίο Β εφαρμόζεται έτσι ώστε να ταιριάζει με τη συχνότητα των ιόντων κυκλοτρονίου. Αυτό επιτυγχάνεται πράγματι με το κύμα Schumann 7,83 Hz του γεωμαγνητικού πεδίου.
Μια τέτοια απομάκρυνση των ιόντων από τις τροχιές των κυκλοτρονίων τους παράγει με τη σειρά της, λόγω της διατήρησης της στροφορμής (γωνιακής ορμής), μια αντίθετη περιστροφή του πλάσματος των σχεδόν ελεύθερων ηλεκτρονίων στα CD, η συχνότητα της οποίας εξαρτάται, φυσικά, από τον αριθμό των εμπλεκόμενων ιόντων, δηλαδή από τη συγκέντρωσή τους, η οποία είναι επομένως η μόνη σημαντική μεταβλητή.
Το φαινόμενο εμφανίζεται με τον ίδιο τρόπο σε όλα τα CD του συστήματος, ο αριθμός των οποίων είναι ασήμαντος για αυτό το σκοπό, σε συμφωνία με το σημείο 2 του τμήματος 2.
Κατά συνέπεια, η επαγόμενη με αυτόν τον τρόπο περιστροφή του πλάσματος των σχεδόν ελεύθερων ηλεκτρονίων στα CDs παράγει το ανιχνεύσιμο σήμα EMS.
Μια επιβεβαίωση ότι οι ανιχνεύσιμες συχνότητες παράγονται από μαγνητική δραστηριότητα δίνεται από την άποψη ότι η διέγερση αποτρέπεται από την απορρόφηση του μ-μετάλλου.
Συνοψίζοντας, η παραπάνω ανάλυση ταιριάζει με το ρόλο που παίζει η συχνότητα στα 7,83 Hz στις παρατηρήσεις (βλ. Ενότητα 2).
Είναι ενδιαφέρον να παρατηρήσουμε ότι, εάν τροχιές των κυκλοτρονίων γύρω από το κέλυφος (shell) του νερού (υδάτινο κέλυφος) κορεστούν από ένα είδος ιόντων που δεν ταιριάζει με τους συντονισμούς Σούμαν, η δραστηριότητα του βιολογικού συστήματος θα ανασταλεί.
Η πρόβλεψη αυτή είναι σύμφωνη με τα γεγονότα, καθώς γνωρίζουμε ότι υπάρχουν ιόντα που προάγουν τη βιολογική δραστηριότητα και ιόντα που την αναστέλλουν.
Το παραπάνω συμπέρασμα ισχύει, βέβαια, αν το μοναδικό υπόβαθρο em είναι το φυσικό (μορφές Σούμαν) ή ένα τεχνητό υπόβαθρο em με συχνότητες παρόμοιες με αυτές του Σούμαν αλλά αν υπήρχε ένα τεχνητό υπόβαθρο em με διαφορετική κατανομή συχνοτήτων, θα γινόταν μια αναδιάταξη των ευνοϊκών και δυσμενών ειδών ιόντων.
Αυτό το χαρακτηριστικό θα μπορούσε να παρέχει μια λογική για την παρατηρούμενη επίδραση των πεδίων ELF στη φυσιολογική δραστηριότητα.
Παρατηρούμε επίσης ότι η εξάρτηση από τη συγκέντρωση είναι η αιτία της εξάρτησης των συχνοτήτων σήματος από την υδατική αραίωση.
Εφόσον η στροφορμή (γωνιακή ορμή) είναι κβαντισμένη, αναμένεται φυσικά ένα κατώφλι στην τιμή της αραίωσης (συγκέντρωση ιόντων), όπως πράγματι παρατηρήθηκε.
Πιο αναλυτικά, αν υποθέσουμε ότι είμαστε σε θέση να εξάγουμε ν ιόντα ανά CD, τότε το πλάσμα των Ν σχεδόν ελεύθερων ηλεκτρονίων αρχίζει να περιστρέφεται με συχνότητα πολύ μεγαλύτερη από τη συχνότητα των ιόντων κυκλοτρονίου, δεδομένου ότι η μάζα των ηλεκτρονίων είναι πολύ μικρότερη από τη μάζα των ιόντων.
Ένα αντίστοιχο πεδίο συν-συντονισμού εμφανίζεται στο περιβάλλον των περιστρεφόμενων CD και θα μπορούσε να είναι η αιτία μιας εκτεταμένης συνοχής μεταξύ των CD.
Η συχνότητα των CD μειώνεται με την αύξηση της υδατικής αραίωσης. Η ύπαρξη ενός παραθύρου αραιώσεων (βλέπε σημείο 3 στο τμήμα 2) θα μπορούσε να γίνει κατανοητή υποθέτοντας ότι το σήμα που παράγεται από τις χαμηλότερες αραιώσεις θα μπορούσε να έχει συχνότητα υψηλότερη από το διάστημα των τιμών που ανιχνεύονται από τα χρησιμοποιούμενα όργανα.
Αντίθετα, οι υψηλότερες αραιώσεις δεν θα µπορούσαν να παράγουν κανένα σήµα επειδή η συγκέντρωση ιόντων µειώνεται κάτω από το όριο που µπορεί να διεγείρει τα CD.
Σημειώνουμε ότι η συνοχή μεταξύ των CDs σε μια περιοχή μεσοσκοπικής έκτασης, που παράγεται με την παρουσία της ενέργειας διέγερσης σε μια τέτοια περιοχή, μπορεί να στηρίξει τη μη διασπορά της διάδοσης του em πεδίου ακτινοβολίας.
Εδώ παίζει έναν ιδιαίτερο ρόλο η συγκεκριμένη δομή του DNA και το συνεκτικό νερό που το περιβάλλει. Αυτό είναι ένα σημείο που πρέπει να αναλυθεί περαιτέρω με βάση τη συγκεκριμένη χημική δομή των τμημάτων (θραυσμάτων) DNA που χρησιμοποιήθηκαν στα πειράματα.
Μια τόσο σημαντική έρευνα είναι στα μελλοντικά μας σχέδια.
Το πεδίο που σχετίζεται με το παρατηρούμενο EMS, το οποίο δημιουργείται μέσω του εξαιρετικά μη γραμμικού μηχανισμού που περιγράφηκε παραπάνω, μπορεί επίσης να προκαλέσει τη συνοχή μεταξύ των CDs σε ένα δεύτερο σωλήνα που περιέχει καθαρό νερό, όπου επιτρέπεται η διάδοσή του υπό τις συνθήκες που έχουν οριστεί στα πειράματα που περιγράφονται στο υποτμήμα 2.1.
Το φαινόμενο αυτό είναι παρόμοιο με το φαινόμενο εγγύτητας που παρατηρείται σε δύο υπεραγώγιμα δείγματα ή στις συστοιχίες επαφών Josephson, με το οποίο τα δείγματα ή οι επαφές περιέρχονται σε καθεστώς κλειδώματος φάσης. Στην υπό μελέτη περίπτωση, ένα τέτοιο καθεστώς κλειδώματος φάσης εκδηλώνεται στην παρατηρούμενη μετάδοση του διεγερμένου em πεδίου των μικροδομών νερού που περιβάλλουν το DNA στον πρώτο σωλήνα.
Μόλις τα CD του νερού του δεύτερου σωλήνα διεγερθούν από το em πεδίο που προέρχεται από τον πρώτο σωλήνα, το DNA κατασκευάζεται σύμφωνα με τη διαδικασία που προτείνεται στο σημείο δ της παρούσας ενότητας. Το κλείδωμα φάσης στη συγκεκριμένη συχνότητα του em πεδίου που διαδίδεται στον αρχικό σωλήνα DNA αντικατοπτρίζεται σαφώς στην ιδιαιτερότητα των επαγόμενων μικροδομών νερού από τις οποίες αναδημιουργείται η αρχική αλληλουχία DNA (98 % ταυτόσημη).
Από την άλλη πλευρά, η παρατηρούμενη υψηλή αναπαραγωγιμότητα του πειράματος αυτού βρίσκει την εξήγησή της στην υψηλή σταθερότητα των συνεκτικών δομών (CDs και συνεκτικές συστάδες CDs) που τίθενται σε λειτουργία. Πρόσφατα πραγµατοποιήθηκε µια σειρά πειραμάτων µε στόχο την επαλήθευση της ορθότητας του περιγραφόμενου θεωρητικού σχήματος.
Τέλος, παρατηρούμε ότι, στο παρόν στάδιο της θεωρητικής ανάλυσης, η συζήτησή μας μπορεί να μας οδηγήσει μόνο σε ποιοτική συμφωνία με τα χαρακτηριστικά που παρατηρήθηκαν στα πειράματα.
Για μια ποσοτική προσαρμογή πρέπει να εισαγάγουμε συγκεκριμένα μοντέλα στο πλαίσιο του γενικού θεωρητικού σχήματος που απεικονίζεται παραπάνω. Το αφήνουμε αυτό για μια μελλοντική εργασία.
4. Συμπεράσματα και ιατρικές εφαρμογές
Στην παρούσα εργασία περιγράψαμε τα πειράματα που έδειξαν μια νέα ιδιότητα του DNA και την επαγωγή ηλεκτρομαγνητικών κυμάτων σε αραιώσεις νερού. Παρουσιάσαμε εν συντομία το θεωρητικό σχήμα που μπορεί να εξηγήσει ποιοτικά τα χαρακτηριστικά που παρατηρήθηκαν σε αυτά τα πειράματα.
Παρατηρούμε ότι είναι δυνατόν να ανιχνευθεί το ίδιο EMS από το πλάσμα ασθενών που πάσχουν από διάφορες λοιμώξεις και χρόνιες ασθένειες. Το πλάσμα πρέπει να διατηρείται φρέσκο και μη κατεψυγμένο.
Εάν το πλάσμα καταψυχθεί στους -70 ◦C, τότε πρέπει να εξαχθεί το DNA προκειμένου να ανακτηθούν τα σήματα. Το DNA μπορεί επίσης να εξαχθεί από βιοψίες ιστών. Ο κατάλογος των ασθενειών στις οποίες έχουν βρεθεί EMS (όπως Αλτσχάιμερ, Πάρκινσον, σκλήρυνση κατά πλάκας, διάφορες νευροπάθειες, σύνδρομο χρόνιου Lyme, ρευματοειδής αρθρίτιδα) δείχνει σαφώς ότι η παρουσία τους δεν περιορίζεται σε ασθένειες που είναι γνωστό ότι έχουν μολυσματική προέλευση: Το γεγονός ότι έχουν βρεθεί EMS σε ασθένειες που δεν είναι γνωστό ότι έχουν μολυσματική προέλευση είναι ενδιαφέρον και μας οδηγεί στην αναζήτηση βακτηριακών ή ιογενών παραγόντων σε αυτές τις ασθένειες.
Μια ειδική περίπτωση είναι αυτή του HIV. Έχουν ανιχνευθεί τακτικά σήματα που προέρχονται από αλληλουχίες DNA του HIV στο αίμα ασθενών που υποβάλλονται σε αντιρετροϊκή θεραπεία και ανταποκρίνονται καλά στην εν λόγω θεραπεία με την εξαφάνιση των ιϊκών αντιγράφων RNA στο κυκλοφορούν αίμα.
Αυτό θα έδειχνε ότι το εν λόγω DNA προέρχεται από μια δεξαμενή που δεν είναι προσβάσιμη στην κλασική θεραπεία και όχι από ιικά σωματίδια που κυκλοφορούν στο αίμα. Επιπλέον, όχι μόνο το πλάσμα των ασθενών, αλλά και τμήμα των ερυθρών αιμοσφαιρίων περιέχει DNA που εκπέμπει σήματα.
Αυτό είναι ενδιαφέρον, καθώς τα ερυθρά αιμοσφαίρια δεν περιέχουν κυτταρικό DNA και καθώς ο ιός δεν συνδέεται με τη μεμβράνη των ερυθρών αιμοσφαιρίων. Η πιθανότητα να εμπλέκεται ένα τρίτο στοιχείο βρίσκεται υπό διερεύνηση.
Έχει διατυπωθεί η υπόθεση [2] ότι η αντιρετροϊκή αγωγή, η οποία περιλαμβάνει αναστολείς της αντίστροφης μεταγραφάσης, επιλέγει η ίδια έναν νέο τρόπο αντιγραφής του ιικού DNA που περιλαμβάνει μία ή περισσότερες κυτταρικές DNA πολυμεράσες.
Όσον αφορά το DNA του M. pirum, προτείνεται ότι τα θραύσματα DNA του HIV και οι νανοδομές τους που υπάρχουν στο αίμα μπορεί να μην προέρχονται από την κυτταρική έκλυση, αλλά, αντίθετα, να αντιπροσωπεύουν κομμάτια συγκεκριμένου μεγέθους ικανά να ανασυνδυαστούν στα κατάλληλα κύτταρα-δέκτες (λεμφοκύτταρα) για να σχηματίσουν ολόκληρο γονιδιωματικό DNA και τελικά να αναγεννήσουν μολυσματικό ιό.
Όποια και αν είναι η προέλευση αυτού του DNA, η εύκολη ανίχνευσή του με ηλεκτρομαγνητικά σήματα μπορεί να το καταστήσει μοναδικό βιοδείκτη για την προσβολή της ιικής δεξαμενής. Η ύπαρξή του στο αίμα με την πιο κλασική τεχνολογία PCR έχει επίσης επιβεβαιωθεί [2].
Συνεπώς, διαθέτει κανείς ένα ισχυρό εργαλείο για τη διενέργεια εξετάσεων νέων τύπων θεραπείας με στόχο την εξάλειψη της λοίμωξης από τον HIV, η οποία δεν έχει επιτευχθεί ποτέ ακόμη, με τις θεραπείες που χρησιμοποιούνται στην πράξη.
Αυτό είναι ιδιαίτερα σημαντικό για τους ασθενείς σε χώρες στις οποίες ο επιπολασμός του HIV είναι πολύ υψηλός (5-10 % σε μεγάλο μέρος της υποσαχάριας Αφρικής).
Σχεδιάζεται η έναρξη κλινικών δοκιμών στη Δυτική και Νότια Αφρική για τη δοκιμή νέων θεραπευτικών ουσιών. Η αποτελεσματικότητά τους θα παρακολουθείται από τη νέα αυτή εξέταση, μαζί με τη βελτίωση των πιο κλασικών παραμέτρων, αξιολογώντας την πλήρη αποκατάσταση του ανοσοποιητικού συστήματος.
Ο στόχος εδώ είναι η εξάλειψη της λοίμωξης από τον ιό HIV, ώστε να μην είναι απαραίτητο οι ασθενείς να υποβάλλονται σε ισόβια θεραπεία με συνδυασμό τοξικών και ακριβών φαρμάκων. Το έργο μας είναι διεπιστημονικό, με τη συμμετοχή βιολόγων, φυσικών και ιατρών. Υπάρχουν βέβαια πολλά άλυτα ερωτήματα που εγείρονται από τα ευρήματά μας, τα οποία αξίζουν περισσότερη δουλειά και περισσότερες αλληλεπιδράσεις.
Η εμπομπή σήματος από το DNA διεγείρεται από φυσικά κύματα 7 Hz που υπάρχουν στη γη. Τα κύματα που παράγονται από τον ανθρώπινο εγκέφαλο είναι επίσης στην περιοχή των 7 Hz. Αυτά είναι ενδιαφέροντα ερωτήματα που πρέπει να τεθούν και ενδεχομένως να απαντηθούν.
Παραπομπές
[1] Montagnier L, Aissa J, Ferris S, Montagnier JL and Lavallee C 2009 Interdiscip. Sci. Υπολογιστής. Life Sci. 1 81–90
[2] Montagnier L, Aissa J, Lavallee C, Mbamy M, Varon J and Chenal H 2009 Interdiscip. Sci. Υπολογιστής. ΖΩΗ Sci. 1 245–253
[3] Montagnier L 2010 Συνέδριο Νόμπελ Lindau
[4] Nickolaenko AP and Hayakawa M 2002 Resonances in the Earth-ionosphere cavity (Dordrecht-Boston Λονδίνο: Kluwer Academic Publishers)
[5] Gibbon DG et al 2010 Science 329 52–56
[6] Preparata G 1995 QED Συνοχή στην ύλη (Σιγκαπούρη: World Scientific)
[7] Arani R, Bono I, Del Giudice E, Preparata G 1995 Int. J. Mod. Phys. B 9 1813–1841
[8] Del Giudice E, Preparata G and Vitiello G 1988 Phys. Αναθ. Lett. 61 1085–1088
[9] Del Giudice E and Vitiello G 2006 Phys. Rev. A 74 022105
[10] Del Giudice E and Tedeschi A 2009 Electr. Biol. Med. 26 48–54
[11] Del Giudice E, Spinetti PR and Tedeschi A 2010 Water 2 566–586 (online Journal)
[12] Del Giudice E, Pulselli RM and Tiezzi E 2009 Ecol. Model 220 1874–1879
[13] Marchettini N, Del Giudice E, Voeikov and VL and Tiezzi E 2010 J. Theo. Bio. 265 511–516 [14] Voeikov VL, Del Giudice E 2009 Water Journal.org. 1 52–75
[15] Landau LD 1941 J. Physics USSR (Μόσχα) 5 71
[16] Szent-Gyorgyi A 1957 Bioenergetics (Νέα Υόρκη, Νέα Υόρκη: Academic Press Inc)
[17] Zhadin M. N, Novikov VV, Barnes FS and Pergola NF 1998 Bioelectromagn. 19(1) 41–49 [18] Zhadin M and Giuliani L 2006 Electr. Biol. Med. 25(4) 269–280
[19] Del Giudice E, Fleischmann M, Preparata G and Talpo G 2002 Bioelectromagnetics 27 522–530.
[20] Del Giudice E and Giuliani L 2010 Eur. J. του Oncol. Βιβλιοθήκη 5 7–23
[1] Montagnier L, A¨ıssa J, Ferris S, Montagnier J-L and Lavallee C 2009 Interdiscip. Sci. Comput. Life Sci. 1 81–90
[2] Montagnier L, A¨ıssa J, Lavallee C, Mbamy M, Varon J and Chenal H 2009 Interdiscip. Sci. Comput. Life Sci. 1 245–253
[3] Montagnier L 2010 Lindau Nobel Conference
[4] Nickolaenko A P and Hayakawa M 2002 Resonances in the Earth-ionosphere cavity (Dordrecht-Boston- London: Kluwer Academic Publishers)
[5] Gibbon D G et al 2010 Science 329 52–56
[6] Preparata G 1995 QED Coherence in matter (Singapore: World Scientific)
[7] Arani R, Bono I, Del Giudice E, Preparata G 1995 Int. J. Mod. Phys. B 9 1813–1841 [8] Del Giudice E, Preparata G and Vitiello G 1988 Phys. Rev. Lett. 61 1085–1088
[9] Del Giudice E and Vitiello G 2006 Phys. Rev. A 74 022105
[10] Del Giudice E and Tedeschi A 2009 Electr. Biol. Med. 26 48–54
[11] Del Giudice E, Spinetti P R and Tedeschi A 2010 Water 2 566–586 (online Journal) [12] Del Giudice E, Pulselli R.M and Tiezzi E 2009 Ecol. Model 220 1874–1879
[13] Marchettini N, Del Giudice E, Voeikov and V L and Tiezzi E 2010 J. Theo. Bio. 265 511–516 [14] Voeikov V.L, Del Giudice E 2009 Water Journal.org. 1 52–75
[15] Landau L.D 1941 J. Physics USSR (Moscow) 5 71
[16] Szent-Gyorgyi A 1957 Bioenergetics (New York, NY: Academic Press Inc)
[17] Zhadin M. N, Novikov V V, Barnes F S and Pergola N F 1998 Bioelectromagn. 19(1) 41–49 [18] Zhadin M and Giuliani L 2006 Electr. Biol. Med. 25(4) 269–280
[19] Del Giudice E, Fleischmann M, Preparata G and Talpo G 2002 Bioelectromagnetics 27 522–530.
[20] Del Giudice E and Giuliani L 2010 Eur. J. of Oncol. Library 5 7–23
Πηγές:
Εναλλακτικά από την πληρωμένη συνδρομή στο Substack, μπορείτε να κάνετε εφάπαξ ή επαναλαμβανόμενες δωρεές μέσω του Ko-Fi:

Το Substack του Nioland είναι μια έκδοση που υποστηρίζεται από τους αναγνώστες. Για να λαμβάνετε νέες δημοσιεύσεις υποστηρίξτε το έργο μου. Γίνετε συνδρομητές δωρεάν ή επί πληρωμή με πλεονεκτήματα.
Λεπτομέρειες για το κόστος και τα πλεονεκτήματα πληρωμένης συνδρομής (μηνιαία ή ετήσια) εδώ:
Η πληροφόρηση είναι ελευθερία, η άγνοια είναι δουλεία... ένας ενημερωμένος πολίτης δεν ξεγελιέται!
Γίνετε κυνηγός πληροφοριών >> Telegram https://t.me/NIO_LAND


Share this post